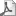|
Одной из важнейщих областей ЦНС, координирующей и контролирующей функции эндокринных желез, является гипоталамус, где локализуются нейросекреторные ядра и центры, принимающие участие в регуляции синтеза и секреции гормонов аденогипофиза. Гипоталамус – область головного мозга, расположенная между перекрестом зрительных нервов, зрительным трактом, внутренним краем ножки головного мозга и сосковидными телами. Гипоталамическая бороздка, идущая от сильвиева водопровода до монроева отверстия, отделяет гипоталамус от зрительного бугра. В гипоталамусе различают три больших зоны: перивентрикулярную, медиальную и латеральную. В свою очередь каждая зона состоит из нескольких ядер. Так, в перивентрикулярной зоне различают одну область и 6 ядер: преоптическую перивентрикулярную область, переднее перивентрикулярное ядро, супрахиазматическое ядро, дорсомедиальное ядро, туберальное магноцеллюлярное ядро, аркуатное или дугообразное ядро (иногда называется инфундибулярным ядром), паравентрикулярно-перивентрикулярное ядро. В срединной зоне гипоталамуса различают медиальную преоптическую область, медиальное преоптическое ядро, переднюю гипоталамическую область, паравентрикулярное ядро, вентромедиальное ядро, перифорникальное ядро, заднюю гипоталамическую область и медиальное мамиллярное (сосочковое) ядро. К латеральной зоне относят латеральную преоптическую область, латеральную гипоталамическую область и супраоптическое ядро.
Экспериментальные исследования с выключением (разрушением) отдельных структур гипоталамуса и нарушением его нервных связей с другими отделами головного мозга позволили установить, что нервный контроль передней доли гипофиза осуществляется двумя механизмами (уровнями регуляции). Первый уровень регуляции реализует так называемая гипофизотропная область гипоталамуса, которая контролирует исходную (базальную) секрецию передней доли гипофиза и нейрогипофизарную секрецию. Второй, более высокий уровень обеспечивается другими гипоталамическими и внегипоталамическими областями мозга (гиппокамп, передний таламус, средний мозг и др.), которые принимают участие в стимуляции или угнетении функции гипофиза. Внегипоталамические структуры мозга осуществляют важный нейроэндокринный контроль деятельности гипофиза и ответственны за суточный ритм секреции гормонов. Средний мозг, гиппокамп и переднемедиальное таламическое ядро участвуют в регуляции секреции АКТГ, гонадотропинов, пролактина, гормона роста. Кроме того, в гипоталамус проецируются также восходящие афферентные и прямые связи из сетчатого образования и среднего мозга, где локализуются дофаминергические и другие клетки, секретирующие различные моноамины. Гипоталамус имеет богатую сеть кровеносных сосудов, которые в области срединного возвышения образуют портальную систему. Наиболее васкуляризированы супраоптическое и паравентрикулярное ядра. Гистологически область срединного возвышения представляет зону контактов, содержащую окончания многочисленных нейронов, локализованных в перечисленных ядрах гипоталамуса, по которым продукты секрета этих нейронов (гипофизотропные гормоны) достигают капилляров воротной (портальной) системы гипофиза. Венозные капилляры портальной системы имеют специальные отверстия (шунты), создающие возможность перехода соединений с достаточной молекулярной массой из крови в периваскулярные пространства срединного возвышения. Гипоталамус, таким образом, является областью, трасформирующей информацию, поступающую по нервным путям из вышележащих отделов нервной системы, посредством изменения уровня нейротрансмиттеров (нейромедиаторов), к которым относятся различные моноамины: адреналин, норадреналин, дофамин, серотонин, ацетилхолин, g-аминомасляная кислота. Стрессовые ситуации и другие факторы приводят к изменению содержания, скорости синтеза и высвобождения моноаминов в гипоталамусе, которые в свою очередь изменяют скорость секреции гипоталамических и гипофизотропных гормонов, что и приводит к соответствующему изменению функциональной активности передней доли гипофиза. Считается, что нейротрасмиттеры (моноамины) регулируют деятельность гипофиза посредством нескольких механизмов: а) участие в синаптической передаче информации, поступающей из лимбической системы мозга, на нейрон, продуцирующий гипофизотропные гормоны (пептиды); б) действие на мембрану гипоталамического нейрона и процесс высвобождения гипофизотропного гормона; в) изменение функциональной активности аксона гипоталамического нейрона в области капилляров портальной (воротной) системы гипофиза с модификацией транспорта гипофизотропного гормона в кровь; г) влияние на клетки передней доли гипофиза с увеличением или угнетением их секреторной активности или модификацией их ответа на действие гипофизотропных гормонов. Таким образом, гипоталамус является местом, где нервные и эндокринные клетки взаимодействуют друг с другом, осуществляя быструю и высокоэффективную передачу информации, необходимой для быстрого ответа со стороны органа, систем и организма в целом с целью обепечения жизнедеятельности организма. Передача информации от клетки к клетке осуществляется химическими мессенджерами (гормоны и моноамины) и электрической активностью. Межклеточные взаимодействия, как показали исследования последних лет, могут осуществляться следующими механизмами: синаптической передачей мессенджера; гормональным механизмом посредством циркулирующих гормонов; паракринным механизмом, т.е. без поступления гормона в кровь, а только в межклеточную жидкость; аутокринным механизмом, т.е. выходом гормона из клетки в межклеточную жидкость и взаимодействие этого гормона с мембранными рецепторами, расположенными на той же клетке. Показано, что норадреналин, соматостатин, дофамин, гонадолиберин, окситоцин, вазопрессин могут выступать в качестве гормонов и секретироваться эндокринными клетками или нейронами, а также выявляться в синапсах нервных клеток и выполнять роль нейротрансмиттеров. Другая группа гормонов – глюкагон, энкефалины, холецистокинин, производные проопиомеланокортина секретируются эндокринными клетками, выполняя гормональную функцию, а также, локализуясь в нервных окончаниях, оказывают нейротрансмиттерное действие. Причем эти два свойства выявляются и у других гормонов аденогипофиза. Тиролиберин и ВИП секретируются нейронами, но выполняют гормональную функцию, а в нервных окончаниях оказывают явное нейротрансмиттерное действие. Влияние ЦНС на функцию гипоталамуса осуществляется не только указанными выше нервными механизмами, но также путем транспорта спинномозговой жидкостью различных гормонов, нейротрансмиттеров и других веществ (эндорфины, энкефалины, вещество Р), вырабатывающихся в различных областях ЦНС и эпифизе. В эпифизе образуются мелатонин и ряд других индолов и полипептидов, модулирующих функцию надпочечников, щитовидной и половых желез. Гормоны эпифиза высвобождаются в спинномозговую жидкость или общий кровоток и действуют различными путями. Так, мелатонин концентрируется в гипоталамусе и среднем мозге и влияет на секрецию гипофизотропных гормонов, изменяя содержание моноаминов и нейромедиаторов. Другие полипептиды эпифиза действуют на образование гипофизотропных пептидов непосредственно. Необходимо отметить, что, помимо нейромедиаторов, в механизмах высвобождения гипофизотропных гормонов принимают обязательное участие ионы К+ и Са2+, простагландины, цАМФ и другие вещества. ПРИНЦИП ОБРАТНОЙ СВЯЗИ В РЕГУЛЯЦИИ ГОРМОНОВОтечественный ученый М.М. Завадовский, изучая закономерности в регуляции деятельности эндокринных желез, впервые в 1933 г. сформулировал принцип “плюс-минус взаимодействие”, получивший в дальнейшем название “принцип обратной связи”. Под обратной связью подразумевается система, в которой конечный продукт деятельности этой системы (например, гормон, нейротрансмиттер и другие вещества) модифицирует или видоизменяет функцию компонентов, составляющих систему, направленную на изменение количества конечного продукта (гормона) или активности системы. Жизнедеятельность всего организма является следствием функционирования многочисленных саморегулируемых систем (выделительная, сердечно-сосудистая, пищеварительная, дыхательная и др.), находящихся в свою очередь под контролем нейроэндокринно-иммунной системы. Все перечисленное представляет, таким образом, комплекс различных саморегулируемых систем, находящихся в определенной степени зависимости и “подчиненности”. Конечный результат или активность системы может модифицироваться двумя путями, а именно путем стимуляции для увеличения количества конечного продукта (гормона) или повышения активности эффекта, или путем угнетения (ингибирования) системы с целью уменьшения количества конечного продукта или активности. Первый путь модифицирования называется положительной, а второй – отрицательной обратной связью. Примером положительной обратной связи является повышение уровня гормона в крови, стимулирующее высвобождение другого гормона (повышение уровня эстрадиола в крови вызывает высвобождение ЛГ в гипофизе), а отрицательной обратной связи, когда повышенный уровень одного гормона угнетает секрецию и высвобождение другого (повышение концентрации тироидных гормонов в крови снижает секрецию ТТГ в гипофизе). Гипоталамо-гипофизарная регуляция осуществляется механизмами, функционирующими по принципу обратной связи, в которых четко выделяются различные уровни взаимодействия (рис. 2).  Рис 2. Уровни функционирования обратной связи.
Под “длинной” цепью обратной связи подразумевается взаимодействие периферической эндокринной железы с гипофизарными и гипоталамическими центрами (не исключено, что и с супрагипоталамическими и другими областями ЦНС) посредством влияния на указанные центры изменяющейся концентрации гормонов в циркулирующей крови. Под “короткой” цепью обратной связи понимают такое взаимодействие, когда повышение гипофизарного тропного гормона (например, АКТГ) модулирует и модифицирует секрецию и высвобождение гипофизотропного гормона (в данном случае кортиколиберина). “Ультракороткая” цепь обратной связи – вид взаимодействия в пределах гипоталамуса, когда высвобождение одного гипофизотропного гормона влияет на процессы секреции и высвобождения другого гипофизотропного гормона. Этот вид обратной связи имеет место в любой эндокринной железе. Так, высвобождение окситоцина или вазопрессина через аксоны этих нейронов и посредством межклеточных взаимодействий (от клетки к клетке) модифицирует активность нейронов, продуцирующих эти гормоны. Другой пример, высвобождение пролактина и его диффузия в межваскулярные пространства приводит к влиянию на соседние лактотрофы с последующим угнетением секреции пролактина. “Длинная” и “короткая” цепи обратной связи функционируют как системы “закрытого” типа, т.е. являются саморегулирующими системами. Однако они отвечают на внутренние и внешние сигналы, изменяя на короткое время принцип саморегуляции (например, при стрессе и др.). Наряду с этим на указанные системы влияют механизмы, поддерживающие биологический циркадный ритм, связанный со сменой дня и ночи. Циркадный ритм представляет собой компонент системы, регулирующий гомеостаз организма и позволяющий адаптироваться к изменяющимся условиям внешней среды. Информация о ритме день-ночь передается в ЦНС с сетчатки глаза на супрахиазматические ядра, которые вместе с эпифизом образуют центральный циркадный механизм – ”биологические часы”. Помимо механизма день-ночь, в деятельности этих “часов” принимают участие другие регуляторы (изменение температуры тела, состояние отдыха, сна и др.). Супрахиазматическим ядрам принадлежит интегрирующая роль в поддержании биологических ритмов. Около 80% клеток супрахиазматических ядер возбуждается при действии ацетилхолина. Попытки изменить ритм деятельности ядер инфузией большого количества серотонина, дофамина, тиролиберина, вещества Р, глицина или g-аминомасляной кислоты оказались неэффективными. Однако в этой области обнаружены некоторые гормоны (вазопрессин, гонадолиберин, вещество Р), которые, несомненно, каким-то образом участвуют в механизмах поддержания биологических ритмов. Секреция многих гормонов (АКТГ, СТГ, глюкокортикоиды и др.) подвержена на протяжении суток значительным колебаниям. На рис. 3 представлен суточный ритм секреции СТГ. Изучение циркадной секреции гормонов имеет большое клиническое значение, так как при некоторых заболеваниях (акромегалия, болезнь Иценко – Кушинга) нарушение суточного ритма секреции гормонов является важным дифференциально-диагностическим признаком, который используется в дифференциации синдромно сходной патологии.  Рис 3. Суточный ритм секреции СТГ.
ГИПОТАЛАМИЧЕСКАЯ РЕГУЛЯЦИЯГипоталамус является той областью ЦНС, которая посредством нейротрансмиттеров, гипоталамических, гипофизотропных гормонов, а также симпатической и парасимпатической частей вегетативной нервной системы интегративно регулирует функциональную активность гипофиза и периферических эндокринных желез.
ГИПОФИЗОТРОПНЫЕ ГОРМОНЫИзвестные в настоящее время гипофизотропные гормоны гипоталамуса делятся на гормоны, усиливающие (высвобождающие, рилизинг-гормоны) и угнетающие (ингибирующие) секрецию и высвобождение (выделение) соответствующих тропных гормонов передней доли гипофиза. Комиссия по биохимической номенклатуре Международного общества чистой и прикладной химии Международного биохимического общества (1974 г.) рекомендовала принять окончание “либерин” в названиях гормонов гипоталамуса, усиливающих высвобождение соответствующих тропных гормонов гипофиза (например, кортиколиберин), и окончание “статин” в названиях гормонов с ингибирующим эффектом (например, соматостатин). Установлено существование следующих гипофизотропных гормонов: 1) гормон, высвобождающий лютеинизирующий и фолликулостимулирующий гормоны – гонадолиберин (люлиберин); 2) кортикотропин-рилизинг-гормон кортиколиберин; 3) соматотропин-рилизинг-гормон – соматолиберин; 4) гормон, угнетающий высвобождение гормона роста – соматостатин; 5) пролактин-рилизинг-гормон – пролактолиберин, функцию которого выполняют, вероятно, тиролиберин и ВИП; 6) гормон, угнетающий высвобождение пролактина – пролактостатин, роль которого выполняет дофамин; 7) тиротропин-рилизинг-гормон – тиролиберин; 8) гормон, высвобождающий меланоцитостимулирующий гормон – меланолиберин; 9) гормон, угнетающий высвобождение меланоцитостимулирующего гормона – меланостатин. Существование двух последних гормонов у человека окончательно не доказано. Гипофизотропные гормоны секретируются нейронами, локализованными в различных областях гипоталамуса. Так, паравентрикулярное ядро гипоталамуса содержит большое количество нейронов, секретирующих тиролиберин и кортиколиберин; дугообразное (аркуатное) ядро содержит нейроны, секретирующие соматолиберин и пролактостатин (дофамин); нейроны, секретирующие соматостатин, располагаются в передней гипоталамической области, а гонадолиберин – в предоптической области. Аксоны перечисленных нейронов заканчиваются в области срединного возвышения гипоталамуса, где начинается портальная система гипофиза, c помощью которой гипоталамус сообщается с передней долей гипофиза. Перечисленные химические медиаторы (гипофизотропные гормоны, моноамины), относящиеся к малым пептидам и биогенным аминам, высвобождаются из гипоталамических нейронов в систему портального кровообращения и, достигая клеток аденогипофиза, модулируют их специфическую активность. Установлено, что нервные терминали (аксоны) нейронов гипоталамуса имеют здесь тесные контакты с первичным капиллярным сплетением, где и происходит высвобождение гипофизотропных гормонов в кровь и их транспорт портальной системой к гипофизу. Концентрация гипофизотропных гормонов в этой системе наивысшая по сравнению с их содержанием в общем кровотоке. Кортиколиберин. Кортитропин-рилизинг-фактор был первым из гипофизотропных гормонов, который был частично охарактеризован еще в 1955 г., однако лишь в 1983 г. W. Vale с сотрудниками представили полную химическую и клиническую его характеристику. Интересно, что для получения 1 мг этого гормона (такое количество необходимо для химической характеристики) исследователи фракционировали 500 тысяч гипоталамусов овцы. В последующие годы кортиколиберин был выделен и из гипоталамуса свиньи, крысы, человека и других животных. Этот пептид состоит из 41 аминокислотного остатка, имеющего молекулярную массу 4758, 14 дальтон. Ген, ответственный за синтез кортиколиберина, локализуется на 8-й хромосоме. Основное количество кортиколиберина локализуется в гипоталамусе, однако он выявляется и в других отделах ЦНС, включая кору головного мозга и различные ядра, где он выполняет роль нейротрансмиттера, координируя ответ на различные стрессовые ситуации. В гипоталамусе кортиколиберин в основнм выявляется в парвоцеллюлярных нейронах паравентрикулярного ядра и нервные аксоны этих нейронов достигают срединного возвышения, где они контактируют с капиллярами портальной системы и с током крови достигают клеток аденогипофиза. Многочисленными исследованиями показано, что адреналэктомия или гипофизэктомия приводит к увеличению содержания кортиколиберина в указанных областях гипоталамуса. Изучение структуры кортиколиберина, полученного из гипоталамуса различных животных, показало, что только кортиколиберин человека и крысы имеет идентичную структуру, которая включает следующую последовательность аминокислот: Ser-Glu-Glu-Pro-Pro-Ile-Ser-Leu-Asp-Leu-Thr-Phe-His-Leu-Leu-ArgGlu-Val-Leu-Glu-Met-Ala-Arg-Ala-Glu-Gln-Leu-Ala-Gln-Gln-Ala-His-SerAsn-Arg-Lys-Leu-Met-Glu-Ile-Ile-NH2. Критическим для сохранения биологической активности гормона является наличие карбоксильного остатка. Исследованиями различных авторов установлено, что фрагмент молекулы, содержащий 15-41 аминокислотный остаток, обладает биологической активностью гормона. Как и другие нейропептиды, кортиколиберин синтезируется из прогормона, включающего 196 аминокислотных остатков. Период полураспада кортиколиберина в плазме составляет около 60 минут. Кортиколиберин селективно увеличивает высвобождение АКТГ и других гормонов, производных общего предшественника-проопиомелпнокортина (ПОМК) (см. ниже). Его влияние на усиление высвобождения АКТГ ингибируется глюкокортикоидами. При этом высокие дозы кортизола уменьшают и даже полностью прерывают его влияние на аденогипофиз. На мембранах кортикотрофов кортиколиберин комплексируется со специфическими высокоаффинными рецепторами, активирует аденилатциклазу, что приводит к повышению внутриклеточного уровня цАМФ, который в свою очередь повышает активность цАМФ-зависимых протеинкиназ. Стимуляция высвобождения АКТГ наблюдается только в присутствии Са2+. Наблюдаемое при этом увеличение уровня внутриклеточного кальция может быть результатом повышения концентрации внутриклеточного цАМФ с последующим фосфорилированием белков кальциевых каналов. Вазопрессин также способен стимулировать высвобождение АКТГ, но для этого требуются дозы, превыщающие в тысячи раз дозы, оказывающие максимальный антидиуретический эффект. Вазопрессин и кортиколиберин оказывают синергическое влияние на секрецию АКТГ. Так, вазопрессин в 2-3 раза усиливает способность кортиколиберина высвобождать АКТГ (прямое потенцирующее действие). Проведенные исследования показали, что в нервных окончаниях срединного возвышения вазапрессин и кортиколиберин выявляются вместе, что свидетельствует о возможной их одновременной секреции при определенных условиях. Потенцирующее действие кортиколиберина на секрецию АКТГ помимо вазопрессина оказывают также адреналин и ангиотензин II. Как известно, вазопрессин осуществляет свое действие через инозитолфосфатную систему, а адреналин и ангиотензин II – через цАМФ. На мембранах клеток передней доли гипофиза выявлены высокоаффинные рецепторы к вазопрессину, которые фармакологически подразделяются на V2 (антидиуретические) и V1 (вазопрессорные) рецепторы. Гипофизотропные нейроны, секретирующие кортиколиберин, локализуются в дугообразном, дорсомедиальном, вентромедиальном ядрах, но наибольшее их количество расположено в паравентрикулярном ядре. Аксоны этих клеток оканчиваются в области срединного возвышения, откуда через портальную систему гипофиза кортиколиберин достигает клеток аденогипофиза.Скорость биосинтеза и высвобождения кортиколиберина модулируется моноаминами. Так, адреналин, норадреналин, серотонин, ацетилхолин, глютамин, ангиотензин II, нейропептид Y и аспартамин стимулируют, а аргинин вазопрессин, g-аминомасляная кислота, вещество Р и опиоидные пептиды угнетают высвобождения кортиколиберина (схема 12). Кроме того, холецистокинин, гастринвысвобождающий пептид, предсердный натрийуретический гормон также способны стимулировать высвобождение АКТГ.  Схема 12. Контроль секреции и высво- бождение кортиколиберина.
КЛН – кортиколиберинсинтезирующий нейрон; 1 – серотонин; 2 – ацетилхолин; 3 – гамма-аминомаслянная кислота; 4 – норадреналин. Сплошные стрелки – стимулирующее влияние, пунктирные – угнетающее влияние на секрецию корти- колиберина. Таким образом, кортиколиберин стимулирует синтез и высвобождение АКТГ посредством цАМФ и системы ФИФ2 (инозитолфосфатная система), являющейся вторичным мессенджером вазопрессина. Повышение концентрации калия деполяризует клеточную мембрану и в присутствии ионов кальция происходит высвобождение АКТГ. Это действие кортиколиберина не требует синтеза белка. Кроме того, кортиколиберин ускоряет биосинтез АКТГ de novo, и это влияние может быть угнетено пуромицином и актиномицином D. Иммуннореактивный кортиколиберин и прокортикотропин-мессенджерная РНК (мРНК) выявляются и в различных органах, например, в мозговом слое надпочечника, печени, поджелудочной железе, спинном мозге, плаценте. Такой внегипоталамический “тканевой” кортиколиберин вызывает более продолжительное, чем гипофизотропный кортиколиберин, высвобождение АКТГ. Исследования показали, что у человека в период беременности уровень иммуннореактивного кортиколиберина увеличивается пропорционально сроку беременности, достигая его пика в период родов. После родов его содержание в крови быстро падает, достигая нормальных значений. Показано, что источником такого кортиколиберина является плацента и плацентарный кортиколиберин модулирует время наступления родовой деятельности посредством изменения чувствительности миометрия к окситоцину. Исследованиями Smith и соавт. (1993) показано, что при этом не происходит одновременного повышения концентрации АКТГ и кортизола в крови, полагая, что это является следствием десенситизации гипофиза. Работами Woods и соавт. (1994) установлено, что отсутствие эффекта от повышенной концентрации кортиколиберина в крови в период беременности связано с одновременным увеличением содержания кортиколиберин-связывающего белка в сыворотке крови. Разработанная ими специальная радиоиммунологическая методика для определения кортиколиберин-связывающего белка позволила не только изучить динамику его секреции в период беременности, но и доказать, что введение кортиколиберина добровольцам-мужчинам приводило к повышению на 46 ± 9% кортиколиберин-связывающего белка, что характерно для его уровня, наблюдаемого в конце беременности. На функцию аденогипофиза оказывает влияние симпатическая нервная система. Принято считать, что это влияние осуществляется через гипоталамус. Как полагает Б.В. Алешин (1971), симпатическим импульсам свойственно хотя бы частично оказывать специфическое действие непосредственно на переднюю долю гипофиза. Соматолиберин. Соматотропинвысвобождающий фактор был выделен из гипоталамуса еще в 1964 г. Однако его химическая структура была установлена лишь в 1980-е годы, когда вначале L. Frohman и соавт. (1981) частично охарактеризовали пептид, изолированный из внегипофизарных опухолей больных, страдающих акромегалией, и обладающий способностью усиливать высвобождение СТГ. Химическая структура соматолиберина была установлена R. Guillemin и соавт. (1981), когда из опухоли поджелудочной железы больного акромегалией был изолирован указанный полипептид. Эти данные были подтверждены J. Rivier и соавт. (1982), а сотрудниками этой же лаборатории была осуществлена экспрессия и клонирование кДНК, кодирующей синтез соматолиберина (Mayo R., 1983). Молекула соматолиберина включает 44 аминокислотных остатка, причем биологическую активность проявляет ее часть с первыми 29 аминокислотными остатками. Пептид с 27 аминокислотными остатками полностью лишен биологической активности. В сыворотке крови и тканях соматолиберин присутствует в различных молекулярных формах. В опухоли из клеток островков поджелудочной железы больных акромегалией обнаружены пептиды, состоящие из 44, 40 и 37 аминокислотных остатков. Ген, ответственный за синтез соматолиберина, локализуется на 20-й хромосоме. Следует указать, что вместо термина соматотропинвысвобождающий гормон или соматолиберин был предложен термин соматокринин, но этот термин в настоящее время практически не используется. У человека основной формой соматолиберина, содержащегося в сыворотке крови, является полипептид, состоящий из 40, а в гипоталамусе – из 44 аминокислотных остатков. Химическая структура молекулы включает следующую последовательность аминокислотных остатков: Tyr-Ala-Asp-Ala-Ile-Phe-Thr-AsnSer-Tyr-Arg-Lys-Val-Leu-Gly-Gln-Leu-Ser-Ala-Arg-Lys-Leu-Leu-Gln-AspIle-Met-Ser-Arg-Gln-Gln-Gly-Glu-Ser-Asn-Gln-Glu-Arg-Gly-Ala-Arg-AlaArg-Leu-NH2. Молекулярная масса соматолиберина-40 составляет 4544,73, а мол. м. соматолиберина-44 -5040, 4 Д. Синтетический соматолиберин у здоровых лиц селективно стимулирует секрецию СТГ, в то время как уровень инсулина, АКТГ, глюкагона и ряда других гормонов в крови при этом не изменяется. Однократное введение соматолиберина приводит к более чем 20-кратному повышению содержания СТГ в крови, которое снижается до исходного уровня в течение 3 ч. Имеются сообщения, что при этом может незначительно изменяться уровень пролактина в крови. В случае инфузии соматолиберина в течение нескольких часов после характерного подъема уровня СТГ выявляется его последующее снижение, что подтверждает его пульсирующую секрецию на протяжении суток (L. Frohman и J. Jansson, 1986). Период полураспада соматолиберина в крови составляет около 7 мин. Исследования показали, что соматолиберин синтезируется в дугообразном (аркуатном) и вентромедиальном ядрах гипоталамуса. Аксоны нейронов указанных ядер оканчиваются в области срединного возвышения. Высвобождение соматолиберина стимулируется серотонином и норадреналином. Стимулирующее влияние лимбической системы на секрецию соматолиберина осуществляется через вентромедиальное ядро гипоталамуса. Связь этого ядра с внегипоталамическими областями в отличие от его связи с областью срединного возвышения является катехоламинергической. Эндорфины стимулируют секрецию СТГ, увеличивая образование соматолиберина. Блокада a-адренергических рецепторов гипоталамуса предупреждает повышение уровня СТГ в сыворотке крови здоровых людей в ответ на гипогликемию, в то время как блокада b-адренергических рецепторов повышает реакцию СТГ на гипогликемию. Механизмы контроля секреции и высвобождения соматолиберина представлены на схеме 13.  Схема 13. Контроль секреции и высво- бождение соматолиберина.
СЛН – соматолиберинпродуцирующий нейрон; 1 – серотонин; 2 – дофамин; 3 – альфа-адренергическое влияние норадреналина; 4 – бета-адренергическое влияние норадреналина. Сплошные стрелки – стимулирующее влияние, пунктирные – угнетающее влияние на секрецию соматолиберина. Показано, что очищенные препараты соматолиберина, комплексируясь с рецепторами, расположенными на мембранах соматотрофов, стимулируют секрецию СТГ посредством активирования аденилатциклазы и повышения количества цАМФ. Наряду с этим активируется фосфатидилинозитоловая система с последующим повышением фосфорилирования различных ферментов гормоно-рецепторного комплекса. Это стимулирующее влияние соматолиберина блокируется соматостатином и усиливается глюкокортикоидами (W. Wehrenberg и соавт., 1983). Соматолиберин широко применяется в клинической практике. Так, проба с внутривенным введением соматолиберина применяется для изучения секреции гормона роста. Чаще используют однократное введение соматолиберина в дозе 50, 100 или 200 мкг (из расчета 1 или 3,3 мкг на 1 кг массы тела). Содержание СТГ в сыворотке крови повышается с 0,7-2,0 до 20-30 нг/мл, а максимальных показателей достигает через 15-30 мин. Соматолиберин применяется и для лечения больных нанизмом, обусловленным первичным поражением гипоталамуса и нарушением вследствие этого секреции СТГ. Соматостатин. При попытках выделить из гипоталамуса овец соматолиберин в лаборатории, руководимой Р. Гелемином, в 1973 г. был получен полипептид, который угнетал высвобождение гормона роста из культуры гипофиза крыс (P. Brazeau и соавт., 1973). В том же году была расшифрована структура соматостатина с такой последовательностью аминокислотных остатков: Ala-Gly-Cys-Lys-Asn-Phe-Phe-Trp-Lys-Thr-Phe-Thr-Ser-Cys. Таким образом, соматостатин является тетрадекапептидом, в 3-м и 14-м положениях содержит два цистеиновых остатка и существует в окисленной и восстановленной формах, причем каждая из них обладает одинаковой биологической активностью. Циклическая форма, по некоторым данным, оказывает более сильное ингибирующее действие на секрецию СТГ и инсулина. L. Pradayrol и соавт. (1980) первыми показали, что в тканях соматостатин присутствует в нескольких формах и, в частности, в форме белка, химическая структура которого включает 28 аминокислотных остатков со следующей последовательностью: Ser-Ala-Asn-Ser-Asn-Pro-Ala-Met-Ala-Pro-Arg-Glu-Arg-Lys-Ala-Gly-Cys-Lys-Asn-Phe-Phe-Trp-Lys-Thr-Phe-Thr-Ser-Cys. Молекулярная масса соматостатина-14 равна 1638, 12 D, а соматостатина-28 -3149 D (дальтон). Ген, ответственный за синтез соматостатина, локализуется на 3-й хромосоме. Обе формы соматостатина являются биологически активными. Цитоиммунохимическими исследованиями показано, что соматостатин-14 выявляется в основном в ЦНС, тогда как соматостатин-28 – в желудочно-кишечном тракте, преимущественно в толстом кишечнике. При использовании иммунолюминесцентного метода было обнаружено, что соматостатин локализуется в нервных окончаниях наружного слоя срединного возвышения и в вентромедиальном ядре, которое считается основным гипоталамическим образованием, осуществляющим регуляцию секреции СТГ. Кроме того, соматостатин выявляется и в области дугообразного ядра, где он присутствует как в клетках и нервных окончаниях, так и в аксонах, проходящих через это ядро. Основные сплетения соматостатинсодержащих нервных волокон располагаются в вентромедиальном и дугообразном ядрах и далее распространяются каудально в вентральные сосцевидные ядра. Установлено также, что нейроны, секретирующие соматостатин, выявляются в паравентрикулярных ядрах. Аксоны этих клеток распространяются латерально и вентрально по направлению к перекресту зрительных нервов, а далее идут каудально к области срединного возвышения. Приведенные выше дугообразное и вентромедиальное ядра получают соматостатинсодержащие нервные волокна из других источников. В частности, нервные окончания, содержащие соматостатин, обнаружены в вентромедиально-дугообразном комплексе, супрахиазматических ядрах и в вентральных премамиллярных ядрах. Эти данные свидетельствуют о том, что соматостатин выполняет функцию не только гормона, но и нейропередатчика или нейромодулятора. Такая возможность подтверждается тем, что соматостатинсодержащие клетки обнаружены в спинальных ганглиях, а нервные волокна, содержащие соматостатин, выявляются в дорсальных рогах спинного мозга, периферических симпатических нейронах, превертебральных ганглиях, нижнем и верхнем мезентериальных узлах. В этой связи интерес представляют работы последних лет, в которых показано, что соматостатин вовлечен в такие важные функции мозга, как сознание, память, двигательная активность, вегетативная и эндокринная регуляция. M-F. Chesselet и соавт. (1995) показали, что соматостатин и его мРНК экспрессируются в полосатом теле мозга, которое вовлечено в двигательную активность и поведенческие реакции организма. Ишемия этих участков мозга увеличивает экспрессию гена соматостатина. И, наоборот, уровень соматостатиновой мРНК снижается после применения галоперидола, антипсихотических препаратов, которые способствуют развитию экстрапирамидных симптомов. Авторы справедливо считают, что выявляемая повышенная экспрессия гена соматостатина при патологических состояниях имеет прямое отношение к появлению при этом различных клиничесих симптомов. Кроме того, перерыв дофаминергической иннервации также приводит к повышению экспрессии гена соматостатина в этой области, что является дополнительным подтверждением участия соматостатина в нарушении двигательных реакций, столь характерных для болезни Паркинсона. Соматостатин оказывает свое биологическое влияние через комплексирование с рецепторами, расположенными на мембранах клеток. В настоящее время различают 5 типов рецепторов к соматостатину, которые неодинаково экспрессируются как в тканях различных областей ЦНС, так и в периферических тканях. Все типы рецепторов экспрессируются в передней доле гипофиза и в гипоталамусе, что указывает на участие этих областей в модуляции секреции гормона роста. M. Berelovitz и соавт. (1995) показали, что сахарный диабет и депривация пищи приводят к уменьшению мРНК рецепторов к соматостатину 1, 2 и 3-го типа в гипофизе примерно на 80% по сравнению с контрольными животными, получающими нормальное питание. При этом количество мРНК рецепторов 4 и 5 типа остается без изменений. Также не изменяется количество мРНК всех видов рецепторов в гипоталамусе. В гипофизе диабетических крыс сниженное количество мРНК рецепторов 1, 2 и 3 типа на 50-80% по сравнению с контрольными животными восстанавливается при проведении инсулинотерапии, но только в отношении мРНК рецепторов 1 типа. Экспрессия мРНК рецепторов 4 типа в гипофизе и всех четырех типов в гипоталамусе при этом остается без изменений. Экспрессия же рецепторов 5 типа в гипофизе, сниженная при диабете на 70% и на 30%, в гмпоталамусе восстанавливается при введении инсулина. Эти исследования четко показывают взаимосвязь количества рецепторов к соматостатину с состоянием обмена веществ. Исходя из структуры и фармакологических свойств рецепторы к соматостатину разделяют на две группы (два семейства): к первой группе относятся рецепторы 2, 3 и 5-го типа, которые раньше по фармакологическим характеристикам относили к соматостатиновым рецепторам 1-го типа; ко второй группе относятся рецепторы 1-го и 4-го типа, ранее называемые соматостатиновыми рецепторами 2-го типа. Таким образом, исследования показали, что соматостатиновые рецепторы относятся к группе с 7 трансмембранными фрагментами и кодируются генами, расположенными на различных хромосомах. Ген рецептора 1-го типа локализуется на 14-й; 2-го типа-на 17-й; 3-го типа-на 22-й; 4-го типа-на 20-й и 5-го типа – на 16-й хромосоме. Рецепторы 1-4-го типа практически эквивалентно комплексируются с соматостатином -14 и 28, тогда как рецепторы 5-го типа почти селективно комплексируются с соматостатином, имеющим структуру, включающую 28 аминокислот. Установлена различная аффинность рецепторов к соматостатину и его аналогам. Так, аналог соматостатина октреотид (SMS 201-995) взаимодействует с рецепторами 2 и 3 типа, очень незначительно с рецепторами 5-го типа. Другой аналог соматостатина – соматулин (BIM 23014) комплексируется с рецепторами 2-го и 5-го типа и лишь незначительно с рецепторами 3-го типа. Изучая экспрессию соматостатиновых рецепторов 1 и 2 типа в гипоталамусе, A. Beaudet и G. Tannenbaum (1995) установили различную их концентрацию в ядрах гипоталамуса (преоптическая область, супрахиазматическое ядро, дугообразное ядро, паравентрикулярное и перивентрикулярное ядро, вентральные премамиллярные ядра), что указывает на участие этих рецепторов в центральной регуляции секреции соматолиберина и соматостатина. Механизмы передачи гормонального сигнала в соматостатиновых рецепторах интенсивно изучаются в последние годы. A. Schonbrunn и соавт. (1995) показали, что соматостатиновые рецепторы в экзокринной части поджелудочной железы осуществляют трансдукцию гормонального сигнала через активирование двух подтипов G-белка: Gia1 и Gia3. Соматостатиновые рецепторы экзокринной части поджелудочной железы относятся к соматостатиновым рецепторам 2-го типа, тогда как в гипофизарных клетках выявляются соматостатиновые рецепторы 1-го и 2-го типа. Взаимодействие соматостатина с клетками гипофиза ведет к увеличению количества соматостатиновых рецепторов независимо от количества вновь синтезированного белка. Работы по идентификации соматостатиновых рецепторов в различных тканях организма неожиданно привели к разработке методов ранней диагностики опухолей, секретирующих различные гормоны. Так, S. Lamberts и соавт. (1995) показали, что рецепторы к соматостатину присутствуют в большинстве опухолей, секретирующих гормоны. Для визуализации рецепторов к соматостатину применяют октреотид, меченный радиоактивным индием. С помощью этого препарата визуализируются в 70-100% опухоли, секретирующие гормон роста, ТТГ, опухоли островков поджелудочной железы, карциноидные опухоли, параганглиомы, феохромоцитомы, медулярный рак щитовидной железы, мелкоклеточный рак легких. Кроме того, менингиомы, рак почек, рак молочной железы, злокачественные лимфомы часто имеют также рецепторы к соматостатину, что позволяет проводить их диагностику с помощью различных сканирующих процедур. Причем исследования, проведенные с использованием соматостатина-14, соматостатина-28 и октреотида, показали различия в количестве связывающих мест к перечисленным гормонам, что указывает на определенные различия в соматостатиновых рецепторах. Помимо этого, для выявления первичной опухоли или ее метастазов во время операции применяется октреотид (аналог соматостатина), меченный радиоактивным йодом. Соматостатин оказывает прямое действие на ЦНС. Он вызывает различные поведенческие, двигательные и электрофизиологические изменения при введении его в гиппокамп, кору головного мозга. Показано, что соматостатин выявляется в d-клетках желудочно-кишечного тракта и поджелудочной железы. В желудке эти клетки располагаются в собственном слое слизистой оболочки антрального отдела, преимущественно в непосредственной близости от клеток, продуцирующих гастрин, что подтверждает влияние соматостатина на секрецию этого гормона. В кишечнике a-клетки выявляются в основном в собственном слое слизистой оболочки; незначительное их количество обнаруживается в криптах. В поджелудочной железе соматостатинсодержащие клетки локализуются по периферии панкреатических островков, располагаясь между a- и b-клетками. Своеобразие распределения d-клеток, а именно их разбросанность среди других эндокринных клеток желудочно-кишечного тракта и поджелудочной железы представляет морфологическую основу для высвобождения и местного действия гормона на соседние клетки-мишени. В этом отношении соматостатин может рассматриваться как аналог других местных гормонов, к которым относятся, в частности, гистамин и серотонин.Кроме того, соматостатин, выявляющийся не только в клетках, где он секретируется, но и в нервных волокнах, в том числе желудочно-кишечного тракта, осуществляет свое действие и через нейрокринные механизмы, т.е. путем высвобождения из нервных окончаний. Соматостатин оказывает влияние на моторную и секреторную функции пищеварительной системы, ее кровообращение и кишечную абсорбцию. Он задерживает эвакуацию желудочного содержимого, угнетая высвобождение мотилина – гормона, стимулирующего моторику желудочно-кишечного тракта, снижает сократитильную активность желчного пузыря путем торможения действия холецистокинина. У накормленных животных соматостатин тормозит высвобождение гастрина и холецистокинина, наблюдаемое после приема пищи. Соматостатин угнетает секрецию поджелудочной железой инсулина и глюкагона. Это действие распространяется на обе фазы высвобождения инсулина, вызванного глюкозой, толбутамидом, глюкагоном или секретином. Из-за короткого периода полураспада соматостатина (около 4 мин) его тормозящее влияние заканчивается вскоре после прекращения инфузии. Угнетая секрецию глюкагона, соматостатин понижает концентрацию глюкозы в периферической крови путем уменьшения выхода ее из печени в воротную вену. Однако после внутривенной нагрузки глюкозой или длительного введения соматостатина вместо ожидаемого снижения уровня глюкозы в крови наблюдается гипергликемия, что объясняется одновременным значительным угнетением секреции инсулина. В настоящее время доказаны следующие влияния соматостатина: 1) ингибирование секреции гастрина, секретина, ТТГ, СТГ, инсулина, глюкагона, мотилина, глицентина, ВИП, ренина; 2) угнетение секреции соляной кислоты и пепсина желудком, уменьшение моторики желудка, ингибирование секреции бикарбонатов и ферментов поджелудочной железой, снижение абсорбции в кишечнике, уменьшение кровотока на всем протяжении желудочно-кишечного тракта, снижение секреции и транспорта желчи. В связи с тормозящим влиянием соматостатина на секрецию СТГ, глюкагона и инсулина высказывались предположения о возможности его использования в лечении сахарного диабета. Оказалось, однако, что соматостатин, угнетая секрецию инсулина, приводит к развитию еще более выраженной инсулиновой недостаточности. Большие надежды возлагались на получение аналогов соматостатина, обладающих исключительным влиянием на ингибирование секреции СТГ и глюкагона и интактным в отношении влияния на секреции инсулина. Полученный аналог соматостатина октреотид обладает рядом преимуществ по сравнению с соматостатином: имеет длительный период полураспада, что позволяет применять его 2-3 раза в сутки; ингибирует патологическую секрецию многих гормонов и может применяться при различных патологических состояниях, включая випому, карциноидный синдром, низидиобластоз, гастриному, соматотропиному (акромегалия). Кроме того, опубликованы данные о применении октреотида при тиротропиноме, желудочном кровотечении, перфорирующей язве желудка, глюкагономе, диарее как у детей, так и у взрослых. Гормон роста усиливает синтез и увеличивает содержание соматостатина в гипоталамусе. Сниженное количество соматостатина в гипоталамусе гипофизэктомированных крыс восстанавливалось до нормы под влиянием экзогенного СТГ. Это указывает на существование положительного механизма обратной связи между СТГ и соматостатином и регуляции секреции СТГ по принципу “короткой”обратной связи. Тиролиберин. Установлено, что высвобождение ТТГ из передней доли гипофиза регулируется гипоталамусом посредством тиротропин-рилизинг-гормона, или тиролиберина. Кстати, именно тиролиберин был первым из гипофизотропных гормонов с установленной химической структурой (C. Bowers и соавт., 1970), которая включает следующую последовательность аминокислот-: pGlu-His-Gly-NH2; мол. м. 362, 42Д. Вскоре после установления структуры тиролиберина он был синтезирован, причем биологическая активность натурального и синтетического препарата оказалась идентичной. Синтез тиролиберина осуществляется посредством посттрансляционного отщепления последнего от большой молекулы предшественника. M. Yamada и соавт. (1990) клонировали и определили структуру человеческой геномной ДНК и гипоталамической кДНК, кодирующей человеческий препротиролиберин.В то же время P. Wu и соавт. (1988), изучая посттрансляционный процессинг предшественника тиролиберина, идентифицировали 3 новых пептида, производных протиролиберина. Установлено, что ген, ответственный за синтез препротиролиберина, локализуется на 3-й хромосоме (M. Yamada и соавт., 1991). Установлено, что экспрессия гена тиролиберина наблюдается как в нормальной, так и в аденоматозной ткани гипофиза. Молекула препротиролиберина человека включает 242 аминокислотных остатка, а аналогичная молекула крысиного препротиролиберина – 255 аминокислотных остатков. Показано, что в синтезе тиролиберина принимает участие тиролиберинсинтетаза, требующая присутствия АТФ и ионов магния. Она была обнаружена в различных областях мозга. Тироксин непосредственно влияет на активность тиролиберинсинтетазы, и синтез тиролиберина осуществляется по принципу отрицательной обратной связи. Цитоиммунохимические исследования показали, что тиролиберин широко представлен в ЦНС и желудочно-кишечном тракте. Нейроны, секретирующие тиролиберин, выявляются в медиальной и парвоцеллюлярной областях паравентрикулярного ядра. Второй по интенсивности содержания тиролиберина является область супрахиазматического преоптического ядра, дорзомедиального ядра и базолатерального гипоталамуса. Аксоны, содержащие тиролиберин, выявляются в дорзомедиальном ядре, медиальной области паравентрикулярного ядра, но наибольшая их концентрация отмечается в области срединного возвышения. Количество тиролиберина, локализованного в гипоталамусе, составляет всего 30-32% от его содержания в мозге. Остальная часть тиролиберина (около 70%) приходится на внегипоталамические области мозга (передний мозг, задняя часть промежуточного мозга, задний мозг, двигательные нейроны спинного мозга, ядра черепных нервов, нейрогипофиз, эпифиз). Тиролиберин, выявляемый в различных частях ЦНС, не является продуктом секреции клеток гипоталамуса. Помимо своей гипофизарной функции, тиролиберин в других областях ЦНС выполняет роль нейропередатчика или нейромодулятора и оказывает в основном стимулирующее влияние. Это проявляется повышением спонтанной двигательной активности, артериального давления, увеличением времени судорожного периода и летальности от стрихнина, увеличением двигательной активности у животных после введения им морфия. Тиролиберин потенцирует возбуждающее действие ацетилхолина на нейроны коры головного мозга, вызывает анорексию, повышает высвобождение норадреналина и дофамина из синаптосом и ускоряет период обмена норадреналина. Кроме того, он участвует в терморегуляции и вызывает гипертермию при введении его в желудочки мозга. Показано, что тиролиберин широко представлен в поджелудочной железе и желудочно-кишечном тракте. В антральном отделе желудка плода человека он выявляется в клетках, содержащих гастрин. Однако у взрослого человека эта колокализация двух гормонов не подтверждается (S. Grasso и соавт., 1992) В противоположность этому тиролиберин выявляется в поджелудочной железе как взрослого, так и плода. При этом в эмбриональном периоде содержание тиролиберина в поджелудочной железе значительно выше. Причем в поджелудочной железе, так же как и в желудке, тиролиберин колокализуется в одной клетке, а именно в b-клетке, содержащей инсулин. У человека назначение тиролиберина снижает скорость абсорбции глюкозы и ксилозы из кишечника (L.O. Dolva и соавт., 1978). Тиролиберин выявляется также в мочеполовых органах (простата, семенные пузырьки и др.). Его наличие выявлено в сетчатой оболочке, плаценте, экстрактах различных опухолей человека. Однако убедительных данных о биологической значимости тиролиберина в этих органах пока нет. Видимо, он оказывает паракринное или нейротрансмиттерное влияние. Механизмы контроля секреции и высвобождения тиролиберина представлены на схеме 14.  Схема 14. Контроль секреции и высвобождение тиролиберина.
ТЛН – тиролиберинсинтезирующий нейрон; 1 – альфа-адренергическое влияние норадреналина; 2 – серотонин; 3 – эндорфины. Сплошные стрелки – стимулирующее влияние, пунктирные – угнетающее влияние на секрецию тиролиберина. Механизм действия тиролиберина включает его взаимодействие со специфическими мембранными рецепторами клеток передней доли гипофиза, ответственных за секрецию ТТГ. Длительное время считалось, что в результате гормонально-рецепторного взаимодействия происходит активация аденилатциклазы с последующим образованием цАМФ, а последний стимулирует секрецию ТТГ. Как показано в последнее время, повышение цАМФ еще недостаточно для стимуляции секреции ТТГ. Действие тиролиберина опосредуется, главным образом, через вторичные мессенджеры фосфатидилинозитоловой системы и последующим фосфорилированием протеинкиназ. Высвобождение ТТГ стимулируется избытком ионов калия и требует обязательного присутствия ионов кальция. Наблюдаемое после взаимодействия тиролиберина с рецепторами повышение внутриклеточного кальция носит двухфазный характер: быстрое повышение внутриклеточного кальция, затем продолжительное плато и второй пик повышения содержания внутриклеточного свободного кальция. Первый пик повышения кальция связан с его высвобождением из внутриклеточного депо под влиянием вторичного мессенджера – инозитолтрифосфата. Открытие кальциевых вольтажзависимых каналов и повышение скорости вхождения кальция из внеклеточной жидкости приводит ко второму пику повышения концентрации внутриклеточного кальция (L. Peizhi и соавт., 1992). Эти и другие авторы справедливо считают, что повышение цАМФ, наблюдаемое после воздействия тиролиберина, является не первичным, а вторичным феноменом, следствием стимулирующего влияния на образование цАМФ вторичных мессенджеров фосфатидилинозитоловой системы. Однако в различных областях ЦНС тиролиберин может первично опосредовать свое действие через образование цАМФ или через инозитолтрифосфат. Количество рецепторов к тиролиберину и их способность к взаимодействию модулируется циркулирующим уровнем тироидных гормонов. Так, у гипотироидных животных количество рецепторов к тиролиберину увеличивается почти в 2 раза по сравнению с контролем. Заместительная терапия тироидными гормонами приводит к уменьшению повышенного количества рецепторов у этих животных (P.M. Hinkle и соавт., 1981). Помимо тироидных гормонов, на процессы взаимодействия тиролиберина с рецепторами большое влияние оказывают простагландины (ПГЕ1). Тиролиберин разрушается в мембранах органов-мишеней, плазме или сыворотке крови под влиянием двух ферментов – пироглутамил аминопептидазы 1-го и 2-го типа (последняя находится в сыворотке крови, а первая связана с мембранами клеток) через трансформацию нескольких соединений. Вначале он трансформируется в кислый тиролиберин, а затем в дипептид – гистидинпролинамид, который подвергается неферментативной циклизации с образованием гистидилпролин дикетопиперазина (циклический гистидил пролин). Показано, что циклический гистидил пролин обладает свойствами пролактолиберина и оказывает дополнительные влияния на ЦНС через повышение уровня цГМФ, что проявляется угнетением аппетита, изменением температуры тела и др. Скорость инактивации тиролиберина в плазме животных увеличивается на фоне предварительного введения трийодтиронина. Тиролиберин накапливается в печени, почках и гипофизе. Период его полураспада составляет около 3-4 мин. У больных, страдающих тиротоксикозом, период полураспада тиролиберина в крови составляет 2 мин, а у больных, страдающих гипотирозом, около 6 мин (E. Iverson, 1991). При снижении функции печени или почек клиренс тиролиберина уменьшается. Несмотря на быструю инактивацию, низкие концентрации тиролиберина определяются в периферической крови. Опыт клиничекого применения тиролиберина показал, что его введение уже в течение первых 5 минут стимулирует высвобождение ТТГ в кровь и последующее повышение уровня тироидных гормрнов. Кроме специфического влияния на ТТГ, тиролиберин увеличивает уровень пролактина в сыворотке крови, проявляя свойства пролактолиберина. Несмотря на многочисленные исследования, окончательного мнения о том, что тиролиберин является физиологическим пролактолиберином, нет. Если порог чувствительности к стимулирующему влиянию тиролиберина на секрецию ТТГ и пролактина одинаков, то акт сосания, стимуляция молочной железы, а также стресс увеличивают содержание пролактина в крови, оставляя интактной концентрацию ТТГ. Более того, циркадный ритм секреции ТТГ и пролактина также не совпадает. На секрецию гормона роста у здоровых людей тиролиберин не влияет, в то время как у больных акромегалией под его влиянием происходит дальнейшее повышение уровня СТГ в сыворотке крови. Это изменение чувствительности к тиролиберину у больных акромегалией, имеющих, как известно, аденому гипофиза, настолько специфично, что проба с ним используется для диагностики и дифференциальной диагностики этого заболевания, а также для оценки эффективности лечения акромегалии. Тиролиберин применяется также для оценки резервов ТТГ и пролактина в гипофизе и дифференциальной диагностики вторичного и третичного гипотироза (см. ”Гипотироз”). При диффузном токсическом зобе уровень ТТГ в сыворотке крови снижен и не изменяется после введения тиролиберина. Кроме того, тиролиберин применяется в неврологической практике. Имеется достаточное количество работ о его положительном влиянии при амиотрофическом латеральном склерозе, дегенерации мозжечка, психической депрессии. Особый интерес представляют работы последних лет, в которых показано положительное влияние тиролиберина в лечении респираторного дистресс-синдрома у новорожденных. Патогенез этого синдрома связан с недостаточным созреванием сурфактантной системы легких, наблюдаемым у недоношенных новорожденных. Назначение тиролиберина или его сочетание с глюкокортикоидами достоверно улучшает отношение лецитин/сфингомиэлин в амниотической жидкости, которое является индексом созревания легочной ткани плода (W. Morales и соавт., 1989; R. Ballard и соавт., 1992). Гонадолиберин. Известно, что репродуктивная функция организма регулируется гипофизом (посредством ФСГ, ЛГ и пролактина), а функция последнего находится под контролем ЦНС, в том числе гипоталамуса. Разрушение гипоталамуса при интактном гипофизе и полной сохранности его кровоснабжения приводит к атрофии гонад и полностью прекращает половое развитие животных. Ранее считалось, что в гипоталамусе секретируется люлиберин, высвобождающий ЛГ, и фоллиберин, высвобождающий ФСГ. Идентификация их структуры и аминокислотного состава (Y. Matsuo и соавт., 1971) показала, что оба пептида имеют одну и ту же последовательность аминокислот: pGlu-His-Trp-Ser-Tyr-Gly-Leu-Arg-Pro-Gly-NH2. Мол. м. -1182, 39. Несмотря на установление химической структуры гонадолиберина была выделена кДНК предшественника человеческого гонадолиберина, состоящего из 92 аминокислот и имеющего молекулярную массу около 10000 дальтон (P. Seeburg и J. Adelman, 1984). Ген, ответственный за синтез гонадолиберина, локализуется на 8-й хромосоме и содержит 4 экзона; 2-й экзон кодирует прогонадолиберин, часть 2-го экзона, 3-й экзон и часть 4-го экзона кодируют так называемый гонадолиберинсочетающийся пептид, состоящий из 56 аминокислот, который, по данным K. Nikolics и соавт. (1985), угнетает секрецию пролактина и стимулирует высвобождение ФСГ и ЛГ. Некоторые авторы не исключают наличия двух гипофизотропных гормонов, регулирующих синтез и высвобождение ФСГ и ЛГ. J. Marshall и R. Kelch (1986) показали, что различное влияние гонадолиберина на секрецию гонадотропинов связано с пульсирующей секрецией гонадолиберина. Быстрочастотная секреция гонадолиберина снижает секрецию обоих гонадотропинов; медленночастотная пульсация секреции гонадолиберина повышает уровень ФСГ относительно ЛГ, а инфузия гонадолиберина с постоянной скоростью ингибирует секрецию как ФСГ, так и ЛГ. В настоящее время ни у кого не вызывает сомнения, что гипоталамическая регуляция ФСГ и ЛГ осуществляется одним гормоном – гонадолиберином, который в высокой концентрации выявляется в области срединного возвышения и organum vasculosum. Нейроны, содержащие гонадолиберин, выявляются в обширной области гипоталамуса от перекреста зрительных нервов до супраоптического ядра, но наибольшее их количество локализуется в медиобазальном гипоталамусе. Их аксоны проецируются в область срединного возвышения, где, как уже сказано, концентрация гонадотропина очень высока. В области вентромедиального и дугообразного ядер концентрация гонадолиберина ниже. У человека наибольшее количество нейронов, содержащих гонадолиберин, локализуется в области медиобазального гипоталамуса между третьим желудочком и срединным возвышением (J. L. King и соавт., 1985). В эмбриональном периоде нейроны, продуцирующие гонадолиберин, находятся вместе с нейронами обонятельного нерва, которые локализуются в области, из которой в последующем образуется верхняя челюсть. Они затем мигрируют через носовую перегородку, достигая переднего мозга, где и локализуются в последующем вместе с нервными нейронами. Нарушение в эмбриональном периоде этой миграции нейронов приводит к развитию синдрома Каллмана, для которого характерен гипогонадизм в сочетании с аносмией (см. ”Заболевания половых желез”). Нейроны, содержащие гонадолиберин, локализуются и в других областях ЦНС, ответственных за эмоциональное и половое поведение. В регуляции секреции гонадолиберина участвуют половые гормоны и моноамины. Так, опиоидные пептиды и g-аминомасляная кислота угнетают, а серотонин, адреналин, норадреналин, нейропептид Y, ангиотензин II и ацетилхолин стимулируют высвобождение гонадолиберина. Секретируемый нейронами гонадолиберин транспортируется аксонами этих клеток к синапсам срединного возвышения со скоростью 1-3 мм/ч. Гипоталамус содержит около 20-85 нг гонадолиберина и последний высвобождается пульсирующим способом. Полагают, что в регуляции пульсирующей секреции гонадолиберина участвуют центральные восходящие a-1-адренергические волокна. В регуляцию пульсирующей активности вовлечены также b-эндорфин, метэнкефалин и динорфин, а также вещество P, нейропептид Y и ВИП. С каждым пульсом из гипоталамуса высвобождается в среднем 65-90 пг гонадолиберина. Из гипоталамуса гонадолиберин портальной системой гипофиза транспортируется в гипофиз, где связывается плазматическими мембранами клеток передней доли. Рецептор гонадолиберина состоит из 327 аминокислот и относится к рецепторам, имеющим 7 трансмембранных фрагментов. Гонадолиберин после взаимодействия с соответствующим рецептором подвергается интернализации, транслокации к лизосомам, где он подвергается под влиянием лизосомальных ферментов деградации. Трансдукция гормонального сигнала после его взаимодействия с рецепторами осуществляется вторичными мессенджерами (инозитолтрифосфат и диацилглицерин), производными полифосфоинозитидной системы, которые стимулируют фосфорилирование белков и мобилизацию кальция, вызывая повышение содержания внутриклеточного кальция, что приводит к высвобождению ЛГ. В гипофизарных клетках S. Izumi и соавт. (1990) идентифицировали 2 типа вольтажзависимых кальциевых каналов (Т-тип – подвергающиеся быстрой активации и инактивации, и Л-тип – медленные дигидропиридинчувствительные каналы, подвергающиеся воздействию вторичных мессенджеров). Активирование протеинкиназы С происходит с участием диацилглицерина и цитозольного свободного кальция. Выше отмечалось, что гонадолиберин секретируется пульсирующим образом (всплески активности продолжаются в течение 8-16 мин и повторяются в интервалах через 37-56 мин) и между количеством высвобожденного гонадолиберина и количеством рецепторов к нему имеются обратные взаимоотношения. Изменение количества рецепторов к гонадолиберину является важным фактором регуляции секреции гонадотропинов. Так, эстрогены и андрогены уменьшают количество рецепторов к гонадолиберину. Рецепторы к гонадолиберину помимо клеток передней доли гипофиза выявлены также в яичниках, где, видимо, гонадолиберин оказывает паракринное влияние. При введении гонадолиберина отмечается его быстрое накопление в печени, почках и гипофизе. Период полураспада гонадолиберина в плазме составляет 5-7 минут. В процессах деградации гонадолиберина участвуют 3 ферментативных системы (пептидазы, эндопептидазы). Пролактостатин. Известно, что ЦНС млекопитающих участвует в регуляции секреции пролактина, которая подвержена различным влияниям (акт сосания, эмоциональный стресс и др.). В противоположность действию на другие гормоны передней доли гипофиза в отношении секреции пролактина гипоталамус оказывает тоническое тормозящее влияние. Перерезка ножки гипофиза, т.е. перерыв связи между гипоталамусом и аденогипофизом, приводит к повышению высвобождения пролактина. В 1974-1977 гг. в лаборатории, руководимой А. Шелли, из гипоталамуса свиньи были получены высокоочищенные фракции пролактостатина, которые, однако, содержали до 15% норадреналина и 2% дофамина. Эти данные позволили предположить, что способность названных препаратов угнетать высвобождение пролактина в определенной степени обусловлена присутствием норадреналина, поскольку катехоламины, как изолированные из гипоталамической ткани, так и синтетические, тоже угнетают высвобождение пролактина. Далее из высокоочищенных гипоталамических экстрактов была изолирована g-аминомасляная кислота, которая, как и пролактостатин, угнетала высвобождение пролактина. Таким образом, катехоламины, дофамин и g-аминомасляная кислота, которые постоянно выявляются в гипоталамических высокоочищенных фракциях пролактостатина, оказались способны оказывать такое же действие, как и сам пролактостатин. Многичисленные исследования, проведенные в последние годы, позволили усомниться в существовании пролактостатина. Ряд исследователей считают, что гипоталамический фактор, угнетающий секрецию пролактина, есть не что иное, как дофамин. В то же время не исключена полностью возможность, что дофамин все-таки стимулирует секрецию и высвобождение реально существующего пролактостатина. Введение дофамина в полость 3-го желудочка вызывает повышение уровня пролактостатической активности плазмы крови, взятой из портальной системы гипофиза, при одновременном уменьшении концентрации пролактина в периферическом русле. Вещества, стимулирующие специфические дофаминовые рецепторы, т.е. агонисты дофамина (апоморфин, l-дофа, алкалоиды спорыньи- 2-a-бромэргокриптин или бромокриптин), также угнетают высвобождение пролактина. Дофамин синтезируется тубероинфундибулярными нейронами, расположенными в области дугообразного и перивентрикулярного ядер медиобазальной области гипоталамуса. Аксоны этих нейронов заканчиваются на сосудах портальной системы гипофиза в области срединного возвышения. Секретируемый нейронами дофамин поступает с кровотоком к клеткам передней доли гипофиза и активирует механизм, ингибирующий высвобождение пролактина. Концентрация дофамина в портальной системе гипофиза достаточна для угнетения высвобождения пролактина. После назначения l-дофа, которая в мозге и в периферических тканях конвертируется в дофамин, приводит к быстрому снижению уровня пролактина в крови как у здоровых лиц, так и у больных с различными формами гиперпролактинемии. Дофамин оказывает свое ингибирующее действие на лактотрофы посредством специфических рецепторов. Он угнетает не только высвобождение, но и биосинтез пролактина. Это ингибирующее действие опосредуется через снижение образования цАМФ, синтеза фосфоинозитола, арахидоновой кислоты и обмена фосфолипидов. Помимо цАМФ, вторичные мессенджеры полифосфоинозидной системы участвуют в механизме действия дофамина. Дофамин угнетает также деление клеток и синтез ДНК, что сопровождается исчезновением секреторных гранул в пролактотрофах и стимулированием процессов кринофагии. Таким образом, в настоящее время считают, что роль пролактостатина в организме выполняет дофамин. Это подтверждает и клиническая практика, показавшая успешное применение для лечения гиперпролактинемии агонистов дофамина (парлодел, лизурид и др.). Пролактолиберин (тиролиберин). Из экстрактов гипоталамуса различных животных (крысы, птицы и др.) изолированы фракции, обладающие способностью высвобождать пролактин. Исследования, проведенные после получения тиролиберина, показали, что он также обладает способностью стимулировать высвобождение пролактина. В этой связи высказывались предположения, что тиролиберин и пролактолиберин являются одним и тем же веществом. Однако в результате изучения суточной секреции пролактина и ТТГ было установлено, что ритм секреции этих двух гормонов различен и пики повышения их уровня в крови не совпадают. Кроме того, акт сосания, который стимулирует высвобождение пролактина до очень высоких цифр, не сопровождается одновременным повышением уровня ТТГ в сыворотке крови. С другой стороны, при первичном гипотирозе, при котором уровень тиролиберина и ТТГ повышен, часто наблюдается или увеличение содержания пролактина в сыворотке крови, или его повышенный ответ на стимуляцию при нормальной исходной концентрации. Можно считать, что в этих случаях снижение уровня тироидных гормонов в крови приводит к увеличению чувствительности пролактотрофов к тиролиберину. Наряду с этим существует предположение, что гипотироз сопровождается недостаточной секрецией пролактостатина, чем и объясняется развитие гиперпролактинемии. Серотонинергический механизм играет большую роль в процессах высвобождения пролактина. Введение серотонина или мелатонина в 3-й желудочек мозга приводит к гиперпролактинемии, в то время как угнетение синтеза серотонина парахлорфенилаланином блокирует высвобождение пролактина в ответ на физиологическую стимуляцию. Секрецию пролактостатина связывают с латеральными туберальными ядрами, а пролактолиберина – с передней гипоталамической областью и областью срединного таламуса. Исследованиями, проведенными в последние годы (N. Ben-Jonathan, 1990; B. Arey и M. Freeman, 1990) показано, что в гипоталамусе имеется еще ряд веществ, обладающих активностью пролактолиберина: ВИП, окситоцин, вазопрессин, пептид-гистидин-изолейцин-27 или PHI-27. Показано, что ВИП стимулирует непосредственно секрецию пролактина гипофизом при его добавлении в инкубационную среду или in vivo. Концентрация ВИП в портальной системе гипофиза достаточна для стимуляции высвобождения пролактина. Серотонин оказывает влияние на секрецию пролактина через стимуляцию высвобождения ВИП. Помимо ВИП, кандидатом пролактолиберина является пептид-гистидин-изолейцин, который имеет структурную гомологию с ВИП и колокализуется с кортиколиберином в нейронах паравентрикулярного ядра. Действие гипоталамических и гипофизотропных гормонов, кроме изменения активности аденилатциклазы, объясняется изменением электрического потенциала клеточной мембраны. Деполяризация ее проводит к высвобождению гормона, тогда как гиперполяризация – к угнетению этого процесса. Следует отметить, что пролактотрофы отличаются от других клеток передней доли гипофиза тем, что деполяризация клеточной мембраны, а следовательно, высвобождение пролактина происходит спонтанно, в то время как состояние гиперполяризации, а значит, и угнетение высвобождения гормона поддерживается пролактостатином. Считается, что рилизинг-гормоны, взаимодействуя с рецепторами, расположенными на клеточной мембране, повышают проницаемость клеточной мембраны для кальция и увеличивают количество внутриклеточного кальция, приводят к высвобождению гормона из гранул. Пролактостатин свое угнетающее действие на высвобождение пролактина также осуществляет путем изменения проницаемости мембраны для кальция. Кроме того, высвобождение пролактина опосредуется активацией цАМФ, мессенджерами полифосфоинозидной системы и угнетается повышением внутриклеточного уровня ПГЕ1. Известно, что пролактин не имеет органа-мишени и пока не доказана регуляция его секреции по принципу “длинной” цепи обратной связи. Однако получены многочисленные данные, что “короткая” отрицательная обратная связь в регуляции секреции пролактина функционирует и регулирующим фактором является уровень секретируемого пролактина. Регуляция осуществляется через активацию дофаминергических нервных окончаний срединного возвышения, повышение концентрации дофамина и пролактостатина. Меланостатин и меланолиберин. На протяжении многих лет проводятся исследования по выяснению механизмов гипоталамической регуляции секреции меланоцитостимулирующего гормона. В лаборатории, руководимой A. Schally (1966-1974), путем ферментативного разрушения окситоцина было получено несколько пептидов, из которых пептид Pro-Leu-Gly-NH2 обладал наибольшей меланостатической активностью (меланостатин-1). Из гипоталамуса крупного рогатого скота был выделен другой пептид – Pro-His-Arg-Phe-Gly-NH2, который также угнетал высвобождение МСГ из гипофиза (меланостатин-2). Помимо меланостатина, из гипоталамуса животных был получен гормон, усиливающий высвобождение МСГ-меланолиберин (пентапептид со следующей последовательностью аминокислотных остатков: Cys-Tyr-Ile-Gly-Asn-OH). Незначительные количества этого гормона снижали содержание МСГ в гипофизе и повышали его уровень в крови экспериментальных животных (крыс). В последующем было показано, что меланолибериновая активность средней доли гипофиза крыс связана с a- и b-МСГ, которые кодируются геном, ответственным за синтез проопиомеланокортина. Секреция производных проопиомеланокортина (ПОМК) из средней доли гипофиза угнетается дофаминергическими нервными волокнами дугообразного ядра гипоталамуса. Что касается меланостатина, то его самостоятельное существование подвергается сомнению. Учитывая, что он был выделен из молекулы окситоцина, считают, что именно этому пептиду должны принадлежать биологические эффекты, полученные в предыдущих исследованиях. Окситоцин широко представлен в различных областях ЦНС, где он выполняет роль нейромодулятора и нейротрансмиттера. Считают, что МСГ является наиболее важным гормоном для функции внегипоталамических отделов ЦНС по сравнению с гормонами передней доли гипофиза. У человека в связи с отсутствием средней доли гипофиза самостоятельно не секретируется ни a-, ни b-МСГ, поэтому нет никаких предпосылок и для наличия меланостатина и меланолиберина в гипоталамусе. Однако клиническое испытание меланостатина-1 показало его значительное нейрофизиологическое влияние на поведение, что позволило применить этот пептид для лечения паркинсонизма и психической депрессии. Другие гипофизотропные гормоны. Исследованиями последних лет показано, что в регуляции секреции гормонов гипофиза принимает участие большая группа гормонов, относящихся по своей химической структуре к различным соединениям. К ним относятся: вещество P, гастринвысвобождающий пептид, ВИП, пептид YY, секретин, пептид-гистидин-метионин-27, ПП, нейропептид Y, нейротензин, мотилин, катакальцин, галанин, кальцитонин, ангиотензин-1 и 2, a-предсердный натрийуретический пептид, натрийуретический пептид-32 мозга, октапептид холецистокинина и др. Новые данные получены о d-сонвызывающем пептиде, который был изолирован в 1977 г. из спинномозговой жидкости кролика при электростимляции таламуса. Этот гормон с последовательностью аминокислот Trp-Ala-Gly-Gly-Asp-Ala-Ser-Gly-Glu вовлечен помимо сна в такие функции организма, как толерантность к стрессу, психомоторная деятельность, прекращение приема алкоголя и наркотиков, регуляция температуры тела. Как показали исследования T. Friedman и соавт. (1994), он угнетает базальную и стимулированную кортиколиберином секрецию АКТГ. Дельта-сонвызывающий пептид участвует в регуляции гипоталамо-гипофизарно-надпочечниковой системы и его суточный ритм был подобен АКТГ и кортизолу. Кроме того, суточный ритм гормона тесно связан с температурой тела, что позволило предположить, что d-сонвызывающий пептид участвует в регуляции сна, температуры тела и влияет на функциональную активность аденогипофиза, в частности, модуляции и секреции АКТГ. ГИПОТАЛАМИЧЕСКИЕ НЕЙРОТРАНСМИТТЕРЫЦентральные биоаминергические нейротрансмиттеры и нейромодуляторы принимают участие в регуляции психической деятельности, поведения, а также функциональной активности гипофиза. Эти вещества синтезируются в нейронах, локализованных в гипоталамической и внегипоталамических областях ЦНС. Аксоны нейронов контактируют с капиллярами портальной системы гипофиза в основном в области срединного возвышения и с током крови достигают клеток аденогипофиза. Наиболее важными гипоталамическими нейротрансмиттерами являются дофамин, норадреналин, серотонин, ацетилхолин, g-аминомасляная кислота. Химическая структура перечисленных соединений представлена на схеме 15.  Схема 15. Химическая структура моноаминов.
Синтез и метаболизм нейротрансмиттеров включает следующие этапы: 1) поглощение нейроном специфических аминокислот – предшественников нейротрансмиттеров (триптофан, тирозин, холин, гистидин, глютамин); 2) синтез моноаминов с участием специфических ферментов; 3) хранение синтезированного моноамина в специфических гранулах, где он защищен от разрушения; 4) высвобождение моноамина из гранул в ответ на различные стимулы, которые вызывают деполяризацию нервной мембраны; 5) транспорт моноаминов к тканям-мишеням (портальная система гипофиза или межклеточная жидкость); 6) связывание высвобожденных нейротрансмиттеров рецепторами постсинаптического нейрона или рецептором пептидергического нейрона, секретирующего один из гипоталамических гормонов; 7) метаболизм свободного (не связавшегося с рецептором) нейротрансмиттера или обратное его поглощение и транспорт в гранулы, где он хранится. Нейротрансмиттеры в тканях мозга присутствуют в концентрациях от нанограммов до микрограммов на 1 г. Ацетилхолин как нейротрансмиттер выявляется в 5-10% синапсов ЦНС. В 25-35% синапсов выявляются нейротрансмиттеры, производные глютамина (g-аминомасляная кислота, аспартат, глютамат). Катехоламины, серотонин и гистамин выявляются в 1-2% синапсов. Нейропептиды синтезируются также в рибосомальном аппарате клетки в виде прогормона. Нейропептид, или нейротрансмиттер, как указано выше, накапливается в нервном окончании и по мере его высвобождения, восстановление запаса в нервном синапсе осуществляется процессом обратного его поглощения и поступлением вновь синтезируемого нейропептида. Высвобождение их из нервного окончания происходит, как правило, пульсирующим, а не тоническим способом. Дофаминергические нейроны локализуются от аркуатного и перивентрикулярного ядер до срединного возвышения гипоталамуса. Эта группа нейронов называется дофаминергической туберогипофизарной системой. Другая дофаминергическая система, которая включает область черного вещества (substantia nigra) и другие структуры переднего мозга, участвует в осуществлении экстрапирамидного контроля. Развитие болезни Паркинсона связывают с ее поражением. Третья дофаминергическая система локализуется в пограничных областях гипоталамуса. Две последние дофаминергические системы независимы от первой. Дофамин оказывает непосредственное влияние на клетки аденогипофиза, выделяясь в портальную систему гипофиза, или изменяет его функцию через секрецию гипофизотропных гормонов, или осуществляет свое влияние сразу на обоих уровнях (гипофиз и гипоталамус). Норадренергические нейроны располагаются главным образом в области дна 4-го желудочка (locus coruleus), откуда норадренергические волокна или поднимаются в гипоталамус, средний и передний мозг, или опускаются в спинной мозг. В гипоталамической области норадренергические волокна контактируют с клетками туберогипофизарной системы или нейрогипофизарными нейронами и структурами срединного возвышения. Серотонинергические нейроны локализуются в продолговатом мозге в ядрах срединного шва, а также гипоталамуса (в области срединного возвышения). Показано, что катехоламины (дофамин, адреналин и норадреналин), индоламины (серотонин, гистамин) представлены в стволе мозга и в нисходящих вентральных и дорзальных путях, иннервирующих лимбические и гипоталамические структуры, включая внутренние и наружные слои срединного возвышения. Нейроны, содержащие ацетилхолин, выявляются в супраоптическом ядре и латеральной преоптической области, а холинергические – в тубероинфундибулярной области. Что касается нейронов, продуцирующих g-аминомасляную кислоту, то они локализуются в медиабазальной, медиальной и задней области гипоталамуса. Аксоны всех нейронов могут взаимодействовать между собой и, оканчиваясь нервными терминалами на капиллярах портальной системы в области срединного возвышения, позволяют нейротрансмиттерам высвобождаться в кровь и достигать клеток передней доли гипофиза, участвуя в регуляции аденогипофизарных гормонов.
|